| Euglenid Temporal range: Eocene (53.5Ma) - recent
[1]
| |
|---|---|

| |
| Euglena viridis, by Ehrenberg | |
|
Scientific classification
| |
| Domain: | Eukaryota |
| Clade: | Discoba |
| Superphylum: | Discicristata |
| Phylum: | Euglenozoa |
| Class: |
Euglenida Butschli 1884, emend. Simpson 1997 |
| Major groups | |
| Synonyms | |
| |
Euglenids or euglenoids are one of the best-known groups of flagellates. They are excavate eukaryotes of the phylum Euglenophyta, classified as class Euglenida or Euglenoidea. Euglenids are commonly found in freshwater, especially when it is rich in organic materials, with a few marine and endosymbiotic members. Many euglenids feed by phagocytosis, or strictly by diffusion. A monophyletic group known as Euglenophyceae have chloroplasts and produce their own food through photosynthesis. [2] [3] [4] This group is known to contain the carbohydrate paramylon.
Euglenids split from other Euglenozoa more than a billion years ago. The plastids in all extant photosynthetic species is the result from secondary endosymbiosis between a phagotrophic eukaryovorous euglenid and a Pyramimonas-related green alga. [5] In January 2024, some fossilized forms of euglenid had been found to be mistakenly classified as Pseudoschizaea shells. [6]
Structure

Euglenoids are distinguished mainly by the presence of a type of cell covering called a pellicle. Within its taxon, the pellicle is one of the euglenoids' most diverse morphological features. [7] The pellicle is composed of proteinaceous strips underneath the cell membrane, supported by dorsal and ventral microtubules. This varies from rigid to flexible, and gives the cell its shape, often giving it distinctive striations. In many euglenids, the strips can slide past one another, causing an inching motion called metaboly. Otherwise, they move using their flagella.
Classification
_(1910)_(17950163265).jpg)
1—2. Ascoglena sp. (Euglenales);
3–4. Cryptoglena sp. (idem);
5–9, 14–15, 24–25, 27–29. Trachelomonas spp. (id.);
10. Eutreptia sp. (Eutreptiales);
11, 20. Astasia spp. (Euglenales);
12. Distigma sp. (Eutreptiales);
13. Menoid[i]um sp. (Rhabdomonadales);
16–18. Colacium sp. (Euglenales);
19, 26. Petalomonas spp. (Sphenomonadales);
21. Sphenomonas sp. (id.);
22–23. Euglenopsis sp. (Euglenales);
30. Peranema sp. (Heteronematales)
The first attempt at classifying euglenids was done by Ehrenberg in 1830, when he described the genus Euglena and placed it in the Polygastrica of family Astasiae, containing other creatures of variable body shape and lacking pseudopods or lorica. Later, various biologists described additional characteristics for Euglena and established different classification systems for euglenids based on nutrition modes, the presence and number of flagella, and the degree of metaboly. The 1942 revision by A. Hollande distinguished three groups, Peranemoidées (flexible phagotrophs), Petalomonadinées (rigid phagotrophs) and Euglenidinées (phototrophs), and was widely accepted as the best reflection of the natural relationships between euglenids, adopted by many other authors. [8] Gordon F. Leedale expanded on Hollande's system, establishing six orders ( Eutreptiales, Euglenales, Rhabdomonadales, Sphenomonadales, Heteronematales and Euglenamorphales) and taking into account new data on their physiology and ultrastructure. This scheme endured until 1986, with the sequencing of the SSU rRNA gene from Euglena gracilis. [8]
Euglenids are currently regarded as a highly diverse clade within Euglenozoa, in the eukaryotic supergroup Discoba. [9] They are traditionally organized into three categories based on modes of nutrition: the phototrophs ( Euglenophyceae), the osmotrophs (mainly the 'primary osmotrophs' known as Aphagea), and the phagotrophs, from which the first two groups have evolved. [10] The phagotrophs, although paraphyletic, have historically been classified under the name of Heteronematina. [9]
In addition, euglenids can be divided into inflexible or rigid euglenids, and flexible or metabolic euglenids which are capable of ' metaboly' or 'euglenid motion'. Only those with more than 18 protein strips in their pellicle gain this flexibility. Phylogenetic studies show that various clades of rigid phagotrophic euglenids compose the base of the euglenid tree, namely Petalomonadida and the paraphyletic ' Ploeotiida'. In contrast, all flexible euglenids belong to a monophyletic group known as Spirocuta, which includes Euglenophyceae, Aphagea and various phagotrophs ( Peranemidae, Anisonemidae and Neometanemidae). The current classification of class Euglenida, as a result of these studies, is as follows: [10] [11] [12] [13] [14]
- Euglenida incertae sedis: Atraktomonas, Calycimonas, Dolium, Dylakosoma, Tropidoscyphus, Michajlowastasia, Parastasiella, Dinemula, Paradinemula, Mononema, Ovicola, Naupliicola, Embryocola, Copromonas.
- Order Petalomonadida Cavalier-Smith 1993
- Order "
Ploeotiida" Cavalier-Smith 1993 (paraphyletic)
- Clade Alistosa Lax & Simpson 2021
- Entosiphon Cavalier-Smith, Chao & Vickerman 2016
- Gaulosia Lax, Cho & Keeling 2023
- Clade Karavia Lax, Cho & Keeling 2023
- Chelandium Lax, Cho & Keeling 2023
- Olkasia Lax, Lee, Eglit & Simpson 2019 [15]
- Clade
Spirocuta Cavalier-Smith, Chao & Vickerman 2016
[16] [Helicales Perschke et al. 2017
[11]
- Clade
Anisonemia Cavalier-Smith 2016
- Order
Anisonemida Cavalier-Smith 2016
- Family Anisonemidae Kent 1880 emend. Cavalier-Smith 2016
- Order
Natomonadida Cavalier-Smith 2016
- Suborder
Metanemina Cavalier-Smith 2016
- Family Neometanemidae Cavalier-Smith 2016
- Suborder
Aphagea Cavalier-Smith 1993 emend. Busse & Preisfeld 2003 [Rhabdomonadina Leedale 1967 emend. Cavalier-Smith 1993
[17]
- Family Astasiidae Kent 1884
- Family Distigmidae Hollande 1942
- Suborder
Metanemina Cavalier-Smith 2016
- Order
Anisonemida Cavalier-Smith 2016
- Order
Peranemida Bütschli 1884
- Family Peranemidae Bütschli 1884
- Clade
Euglenophyceae Schoenichen 1925, emend. Marin & Melkonian 2003 [Euglenea Butschli 1884, emend. Busse & Preisfeld 2002]
- Euglenophyceae incertae sedis: Ascoglena, Euglenamorpha, Euglenopsis, Glenoclosteroium, Hegneria, Klebsina, Euglenocapsa.
- Order
Rapazida Cavalier-Smith 2016
- Family Rapazidae Cavalier-Smith 2016
- Order
Eutreptiales Leedale 1967, emend. Marin & Melkonian 2003
- Family Eutreptiaceae Hollande 1942
- Order
Euglenales Leedale 1967, emend. Marin & Melkonian 2003
- Family Phacaceae Kim et al. 2010
- Family Euglenaceae Dujardin 1841, emend. Kim et al. 2010
- Clade
Anisonemia Cavalier-Smith 2016
Nutrition
The classification of euglenids is still variable, as groups are being revised to conform with their molecular phylogeny. Classifications have fallen in line with the traditional groups based on differences in nutrition and number of flagella; these provide a starting point for considering euglenid diversity. Different characteristics of the euglenids' pellicles can provide insight into their modes of movement and nutrition. [18]
As with other Euglenozoa, the primitive mode of nutrition is phagocytosis. Prey such as bacteria and smaller flagellates is ingested through a cytostome, supported by microtubules. These are often packed together to form two or more rods, which function in ingestion, and in Entosiphon form an extendable siphon. Most phagotrophic euglenids have two flagella, one leading and one trailing. The latter is used for gliding along the substrate. In some, such as Peranema, the leading flagellum is rigid and beats only at its tip.
Osmotrophic euglenoids
Osmotrophic euglenids are euglenids which have undergone osmotrophy.
Due to a lack of characteristics that are useful for taxonomical purposes, the origin of osmotrophic euglenids is unclear, though certain morphological characteristics reveal a small fraction of osmotrophic euglenids are derived from phototrophic and phagotrophic ancestors. [19]
A prolonged absence of light or exposure to harmful chemicals may cause atrophy and absorption of the chloroplasts without otherwise harming the organism. A number of species exists where a chloroplast's absence was formerly marked with separate genera such as Astasia (colourless Euglena) and Hyalophacus (colourless Phacus). Due to the lack of a developed cytostome, these forms feed exclusively by osmotrophic absorption.
Reproduction
Although euglenids share several common characteristics with animals, which is why they were originally classified as so, no evidence has been found of euglenids ever using sexual reproduction. This is one of the reasons they could no longer be classified as animals.[ dubious ]
For euglenids to reproduce, asexual reproduction takes place in the form of binary fission, and the cells replicate and divide during mitosis and cytokinesis. This process occurs in a very distinct order. First, the basal bodies and flagella replicate, then the cytostome and microtubules (the feeding apparatus), and finally the nucleus and remaining cytoskeleton. Once this occurs, the organism begins to cleave at the basal bodies, and this cleavage line moves towards the center of the organism until two separate euglenids are evident. [20] Because of the way that this reproduction takes place and the axis of separation, it is called longitudinal cell division or longitudinal binary fission. [21]
Gallery
-
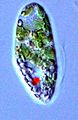 Euglena sp. (Euglenales)
Euglena sp. (Euglenales) -
 Phacus sp. (Euglenales)
Phacus sp. (Euglenales) -
 Trachelomonas sp. (Euglenales)
Trachelomonas sp. (Euglenales) -
 Euglenoid cultures in Petri dishes
Euglenoid cultures in Petri dishes -
 Cell diagram
Cell diagram -
 Astasia sp. (Euglenales)
Astasia sp. (Euglenales) -
 Euglena, Astasia and Phacus spp. (Euglenales)
Euglena, Astasia and Phacus spp. (Euglenales) -
 Euglena, Phacus and Lepocinclis spp. (Euglenales)
Euglena, Phacus and Lepocinclis spp. (Euglenales) -
 Anisonema, Petalomonas, Notosolenus, Scytomonas and Tropidoscyphus spp. (Sphenomonadales); Heteronema, Dinema and Entosiphon spp. (Heteronematales)
Anisonema, Petalomonas, Notosolenus, Scytomonas and Tropidoscyphus spp. (Sphenomonadales); Heteronema, Dinema and Entosiphon spp. (Heteronematales)

.jpg)




_(1910)_(17762559370).jpg)
_(1910)_(17947077272).jpg)
References
- ^ Lee, R.E. (2008). Phycology, 4th edition. Cambridge University Press. ISBN 978-0-521-63883-8.
- ^ Karnkowska, Anna; Bennett, Matthew S.; Triemer, Richard E. (2018). "Dynamic evolution of inverted repeats in Euglenophyta plastid genomes". Scientific Reports. 8 (1): 16071. Bibcode: 2018NatSR...816071K. doi: 10.1038/s41598-018-34457-w. PMC 6207741. PMID 30375469.
- ^ Yoshihisa Hirakawa (2017). Secondary Endosymbioses. Academic Press. p. 323. ISBN 9780128026809.
- ^ "Algaebase :: Subclass: Euglenophycidae". Archived from the original on 2020-07-13. Retrieved 2019-10-27.
- ^ Zakryś, B; Milanowski, R; Karnkowska, A (2017). "Evolutionary Origin of Euglena". Euglena: Biochemistry, Cell and Molecular Biology. Advances in Experimental Medicine and Biology. Vol. 979. pp. 3–17. doi: 10.1007/978-3-319-54910-1_1. ISBN 978-3-319-54908-8. PMID 28429314.
- ^ Cassella, Carly (21 January 2024). "Bizarre Fossils Are Neither Plant Nor Animal, But a 'Weird Fusion' of Life". ScienceAlert. Archived from the original on 22 January 2024. Retrieved 22 January 2024.
- ^ Leander, Brian S.; Farmer, Mark A. (2001-03-01). "Comparative Morphology of the Euglenid Pellicle. II. Diversity of Strip Substructure". Journal of Eukaryotic Microbiology. 48 (2): 202–217. doi: 10.1111/j.1550-7408.2001.tb00304.x. ISSN 1550-7408. PMID 12095109. S2CID 2109559.
- ^
a
b Carlos E. de M. Bicudo; Mariângela Menezes (16 March 2016). "Phylogeny and Classification of Euglenophyceae: A Brief Review". Frontiers in Ecology and Evolution. 4.
doi:
10.3389/FEVO.2016.00017.
ISSN
2296-701X.
Wikidata
Q57898656.
{{ cite journal}}: CS1 maint: unflagged free DOI ( link) - ^ a b Sina M. Adl; David Bass; Christopher E. Lane; et al. (1 January 2019). "Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes". Journal of Eukaryotic Microbiology. 66 (1): 4–119. doi: 10.1111/JEU.12691. ISSN 1066-5234. PMC 6492006. PMID 30257078. Wikidata Q57086550.
- ^ a b Gordon Lax; Alastair G. B. Simpson (16 August 2020). "The Molecular Diversity of Phagotrophic Euglenids Examined Using Single-cell Methods". Protist. 171 (5): 125757. doi: 10.1016/J.PROTIS.2020.125757. ISSN 1434-4610. PMID 33126020. Wikidata Q101127864.
- ^ a b G. Lax; M. Kolisko; Y. Eglit; et al. (June 2021). "Multigene phylogenetics of euglenids based on single-cell transcriptomics of diverse phagotrophs". Molecular Phylogenetics and Evolution. 159: 107088. doi: 10.1016/J.YMPEV.2021.107088. ISSN 1055-7903. Wikidata Q110667805.
- ^ Alexei Y. Kostygov; Anna Karnkowska; Jan Votýpka; Daria Tashyreva; Kacper Maciszewski; Vyacheslav Yurchenko; Julius Lukeš (10 March 2021). "Euglenozoa: taxonomy, diversity and ecology, symbioses and viruses". Open Biology. 11: 200407. doi: 10.1098/RSOB.200407. ISSN 2046-2441. PMC 8061765. PMID 33715388. Wikidata Q125548575.
- ^ Gordon Lax; Anna Cho; Patrick J. Keeling (30 March 2023). "Phylogenomics of novel ploeotid taxa contribute to the backbone of the euglenid tree". Journal of Eukaryotic Microbiology. 70 (4). doi: 10.1111/JEU.12973. ISSN 1066-5234. Wikidata Q123348233.
- ^ Lax G, Keeling PJ (2023). "Molecular phylogenetics of sessile Dolium sedentarium, a petalomonad euglenid". The Journal of Eukaryotic Microbiology. 70 (e12991): e12991. doi: 10.1111/jeu.12991. PMID 37424051.
- ^ Gordon Lax; Won Je Lee; Yana Eglit; Alastair Geoffrey Brinley Simpson (23 March 2019). "Ploeotids Represent Much of the Phylogenetic Diversity of Euglenids". Protist. 170 (2): 233–257. doi: 10.1016/J.PROTIS.2019.03.001. ISSN 1434-4610. PMID 31102975. Wikidata Q92132357.
- ^ Thomas Cavalier-Smith; Ema E Chao; Keith Vickerman (28 August 2016). "New phagotrophic euglenoid species (new genus Decastava; Scytomonas saepesedens; Entosiphon oblongum), Hsp90 introns, and putative euglenoid Hsp90 pre-mRNA insertional editing". European Journal of Protistology. 56: 147–170. doi: 10.1016/J.EJOP.2016.08.002. ISSN 0932-4739. PMID 27718409. Wikidata Q31135651.
- ^ Thomas Cavalier-Smith (15 September 2016). "Higher classification and phylogeny of Euglenozoa". European Journal of Protistology. 56: 250–276. doi: 10.1016/J.EJOP.2016.09.003. ISSN 0932-4739. PMID 27889663. Wikidata Q39151632.
- ^ Leander, Brian Scott (May 2001). "Evolutionary morphology of the euglenid pellicle". University of Georgia Theses and Dissertations.
- ^ Busse, Ingo; Preisfeld, Angelika (14 April 2018). "Systematics of primary osmotrophic euglenids: a molecular approach to the phylogeny of Distigma and Astasia (Euglenozoa)". International Journal of Systematic and Evolutionary Microbiology. 53 (2): 617–624. doi: 10.1099/ijs.0.02295-0. PMID 12710635.
- ^ "Euglenida". tolweb.org. Retrieved 2017-03-30.
- ^ "Reproduction". Euglena. Retrieved 2017-03-31.
Bibliography
- Ciugulea, I.; Triemer, R. E. (2010). A Color Atlas of Photosynthetic Euglenoids. East Lansing, MI: Michigan State University Press. ISBN 9780870138799.
- Leander, B. S.; Triemer, R. E.; Farmer, M. A. (2001). "Character evolution in heterotrophic euglenids". European Journal of Protistology. 37 (3): 337–356. doi: 10.1078/0932-4739-00842. S2CID 4181281.
- Leander, B.S.; Lax, G.; Karnkowska, A.; Simpson, A.G.B. (2017). "Euglenida". In Archibald, J.M.; Simpson, A.G.B.; Slamovits, C. (eds.). Handbook of the Protists. Springer. pp. 1–42. doi: 10.1007/978-3-319-32669-6_13-1. ISBN 978-3-319-32669-6.
- Leedale, G. F. (1978). "Phylogenetic criteria in euglenoid flagellates". BioSystems. 10 (1–2): 183–187. Bibcode: 1978BiSys..10..183L. doi: 10.1016/0303-2647(78)90040-0. PMID 656566.
- Wołowski, K; Hindák, F (2005). Atlas of Euglenophytes. Krakow: VEDA Publishing House of the Slovak Academy of Sciences. ISBN 9788022408363.
External links
-
 Data related to
Euglenoidea at Wikispecies
Data related to
Euglenoidea at Wikispecies - The Euglenoid Project
- Tree of Life: Euglenida
| Authority control databases: National |
|---|